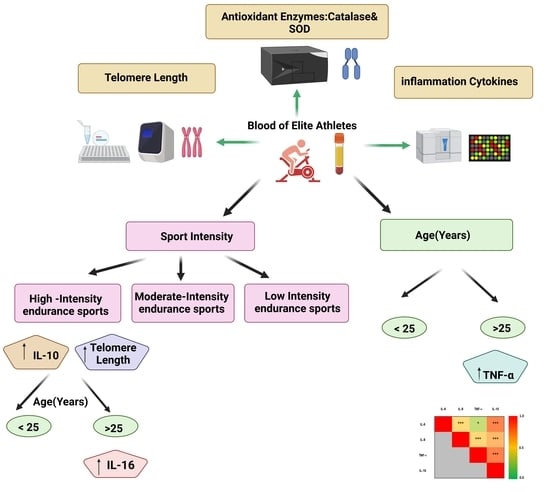
Exercise-associated immune response plays a crucial role in the aging process. The aim of this study is to investigate the effect of sport intensity on cytokine levels, oxidative stress markers and telomere length in aging elite athletes. In this study, 80 blood samples from consenting elite...
Abstract
Exercise-associated immune response plays a crucial role in the aging process. The aim of this study is to investigate the effect of sport intensity on cytokine levels, oxidative stress markers and telomere length in aging elite athletes. In this study, 80 blood samples from consenting elite athletes were collected for anti-doping analysis at an anti-doping laboratory in Italy (FMSI). Participants were divided into three groups according to their sport intensity: low-intensity skills and power sports (LI, n = 18); moderate-intensity mixed soccer players (MI, n = 31); and high-intensity endurance sports (HI, n = 31). Participants were also divided into two age groups: less than 25 (n = 45) and above 25 years old (n = 35). Serum levels of 10 pro and anti-inflammatory cytokines and two antioxidant enzymes were compared in age and sport intensity groups and telomere lengths were measured in their respective blood samples. Tumor necrosis factor-alpha (TNF-α) was the only cytokine showing significantly higher concentration in older athletes, regardless of sport intensity. Interleukin (IL)-10 increased significantly in HI regardless of age group, whereas IL-6 concentration was higher in the older HI athletes. IL-8 showed a significant interaction with sport intensity in different age groups. Overall, significant positive correlations among levels of IL-6, IL-10, IL-8 and TNF-α were identified. The antioxidant catalase activity was positively correlated with levels of TNF-α. Telomere length increased significantly with sport intensity, especially in the younger group. HI had longer telomeres and higher levels of pro- and anti-inflammatory cytokines, suggesting less aging in HI compared to low and moderate counterparts in association with heightened immune response. Investigation of the functional significance of these associations on the health and performance of elite athletes is warranted
So it appears that the intensity of exercise pays a very big part in the ability to live longer. The more intensively the discipline you practice, the slower your cells age. This was show by the increased length of telomers and the higher levels of pro- and anti-inflammatory cytokines. Note intensity also pays a big part in the immune system.
It would be interesting to contrast this study with the effects of protein synthesis and the MTOR pathway. You would have to separate diet from exercise and just look at the exercise factor in longevity.
This would not likely fit the body builder profile as diet is a major factor in muscle growth. Endurance athletes are the ones that appear to benefit the most.
The kinase mammalian target of rapamycin (mTOR) is a central regulator of cell growth and proliferation that integrates inputs from growth factor receptors, nutrient availability, intracellular ATP (adenosine 5'-triphosphate), and a variety of stressors. Since early works in the mid-1990s uncovered the role of mTOR in stimulating protein translation, this kinase has emerged as a rather multifaceted regulator of numerous processes. Whereas mTOR is generally activated by growth- and proliferation-stimulating signals, its activity can be reduced and even suppressed when cells are exposed to a variety of stress conditions. However, cells can also adapt to stress while maintaining their growth capacity and mTOR function. Despite knowledge accumulated on how stress represses mTOR, less is known about mTOR influencing stress responses. In this review, we discuss the capability of mTOR, in particular mTOR complex 1 (mTORC1), to activate stress-responsive transcription factors, and we outline open questions for future investigation.
Keep up to date with news and views on current physiological issues with our quarterly Members’ magazine Physiology News.
www.physoc.org
The stress of exercise
How it tries to break us but ultimately shapes us
Features Mark Burnley
Senior Lecturer, School of Sport and Exercise Sciences, University of Kent, UK The stress of exercise
Muscular exercise is perhaps the most common form of stress that humans experience. Exercise physiology, in turn, is the study of how the body responds to this form of stress. Exercise physiologists use exercise stress experimentally to study physiological control mechanisms. Exercise tests are also used to screen people for cardiovascular, respiratory, and muscular diseases, because a reduced ability to cope with exercise stress is a hallmark of such disorders. Specific disease states frequently produce unique physiological responses to muscular activity, aiding clinical diagnosis. At the other end of the spectrum, regular exercise testing is a common component of athletic training prescription and performance prediction. Understanding exercise stress and how to impose it is the primary mission of the exercise physiologist. Doing so allows us to assess, maintain, or enhance functional capacity in both health and athletic contexts.
Mark Burnley
Defining the stress of exercise
It would be impossible to comprehensively review all forms of ‘exercise-related stress’ in this article. It will instead focus on acute physiological responses to exercise and the chronic adaptations to repeated bouts of exercise (i.e. physical training). The influence of other stressors relevant to exercise (chiefly environmental stress) are mentioned only briefly.
A textbook definition of ‘exercise stress’ does not exist, largely because ‘stress’ is used as a catch-all term for anything that results in a physiological response to muscular activity. As a catch-all definition, this is not a bad start. To be a little more precise, we could define it as ‘Any muscular activity that challenges homeostasis and produces a measurable physiological response’. Writing the previous sentence involved muscle activity, but it would not qualify as stress because it would not have resulted in a measurable physiological response. Rising from the chair to make a cup of tea, however, would qualify, given the contraction of the lower limbs and the changes in blood pressure that occur when standing up. This is one reason why the ‘sit-to-stand’ test is used in the functional assessment of older adults. To begin to understand exercise stress, we must first understand the stimulus causing it before considering the resulting physiological responses.
Muscle activity: the stimulus
Muscle activity comes in many forms, and is used for many purposes. This is one of the reasons exercise is a stressor: the neuromuscular system needs to be a jack of all trades, and few humans are phenotypic specialists. Those that are often become athletes. But even the action of activating a muscle produces a myriad of different stressors which have both acute and chronic effects. Take, for example, the characterisation of muscle action as concentric, isometric, and/or eccentric. In many human activities, all three contraction types take place at various parts of a movement. Each imposes its own unique mechanical and physiological stress on the body. Isometric actions, for example, in which the muscle attempts to shorten, but does not succeed beyond the elastic nature of the muscle–tendon unit, can occlude both venous and (if sufficiently forceful) arterial blood flow, both of which will serve to amplify the metabolic and cardiovascular stress of exercise. Eccentric actions, in which the muscle is lengthened under load, can occur at considerably higher forces than those obtained isometrically, and in such cases the muscle is damaged, leading to an acute loss of maximal force-generating capacity and an inflammatory response over subsequent hours and days. This results in the soreness known colloquially as ‘DOMS’ – delayed-onset muscle soreness.
Concentric actions, in which the muscle truly contracts, are characteristic of heavy resistance exercise using free weights and other machinery. Such exercise can produce rapid fatigue but also influences respiratory and cardiovascular control as the abdominal muscles are forced to contract to brace the thorax. Holding one’s breath in conjunction with bracing the abdomen leads to the classic Valsalva manoeuvre, in which venous return and thus cardiac output are substantially reduced, resulting in light-headedness.
Locomotory activity, in which a substantial fraction of the skeletal muscle is activated (such as running, cycling, rowing, and swimming), involves low-force dynamic muscle actions which can, at various points, be concentric, eccentric, or isometric. Such exercises place a potent stress on the respiratory and cardiovascular systems, because even in untrained individuals the muscle mitochondrial capacity for oxygen utilisation far outstrips the cardiovascular system’s ability to transport oxygen from the lungs. Similarly, the muscle’s ability to produce force far outstrips the mitochondria’s ability to provide a sustainable energy supply for continued force production. These two factors underlie the direct stress which exercise produces, and both form the basis of exercise training for endurance and strength.
Absolute and relative exercise stress
Although exercise intensity is the most important component of exercise stress, a complete definition would require us to consider the duration of exercise and thus the exercise ‘volume’. This is important because even mild exercise can result in significant levels of stress if continued for long enough, as anybody who has hiked anywhere can attest. A distinction also needs to be made between the absolute and relative stress that exercise produces. This is easiest to conceptualise with respect to exercise intensity. The absolute intensity can be expressed simply as the amount of force, speed, or power output the exercise requires. Intensity can also be expressed relative to an individual’s physiological capabilities: as a percentage of maximal heart rate, maximal oxygen uptake, maximal power, or, in the case of maximal contractile force, as a percentage of the maximal voluntary contraction. It is then straightforward to compare individuals’ capabilities in terms of the percentage requirement of an absolute intensity. For example, in cycling, a power output of 250 W might require 80% of maximal oxygen uptake in one individual but only 60% in another. Relative intensity is usually the more meaningful physiological measure of exercise stress, as detailed in ‘Acute responses to exercise stress’ below.
There are situations in which the absolute stress a given task places on an individual is important. Attempts to break a world record in any athletic event require the athlete to achieve a given speed for a known duration. The athlete’s physiological capabilities can then be determined under laboratory conditions to determine if an attempt is feasible. In the uniformed services (police, fire service, and the various branches of the military), there are accepted minimum fitness standards required of all recruits. These standards become increasingly rigorous as the demands of the service increase. In the extreme, the demands placed on special forces soldiers are such that recruits are sometimes placed at significant physical risk. It should be noted, however, that death during vigorous exercise is extremely rare, amounting to one death per ~1.5 million episodes (Albert et al., 2000). However, exercising in the heat or at altitude considerably increases the cardiovascular and/or thermal strain, and thus the acute risk, associated with exercise.
Acute responses to exercise stress
Appreciating the stress of exercise begins with an appreciation of the manner in which energy is transferred within skeletal muscle. Very small quantities of ATP are stored in the muscle. So small, in fact, that a maximal contraction lasting less than 2 seconds could exhaust the muscle’s supply if no other energy sources were available to buffer its loss. One of the triumphs of 20th century biochemistry and physiology was to identify both the metabolic pathways supporting ATP homeostasis and, crucially, how they work in vivo. The fall in ATP that occurs during exercise is buffered by substrate-level phosphorylation (phosphorylcreatine breakdown and glycolysis leading to lactate production) and oxidative phosphorylation. These processes are interlinked via the ‘phosphorylcreatine shuttle’, in which the fall in phosphorylcreatine and the resultant alterations in phosphorylation potential drives mitochondrial respiration. This control mechanism can now be studied non-invasively using magnetic resonance spectroscopy and breath-by-breath measurements of oxygen uptake. Both measurements can be used to infer the stress produced by a given bout of exercise.
A muscle or muscle group that is coping with exercise-induced stress is one that can achieve a steady-state metabolic and respiratory profile (Fig. 1), in which there is no systematic change in muscle metabolites or muscle oxygen utilisation. How large the adjustment is once a steady state is reached (in this context, the rise in oxygen uptake or fall in phosphorylcreatine) gives information about the absolute magnitude of the stressor. However, it is only non-steady- state behaviour that provides information about the relative exercise stress. During ‘moderate’ constant-load exercise (performed below the so-called lactate threshold), a steady state is reached within 2-3 minutes, the metabolic disturbance is small, and exercise can be maintained for several hours if the participant is motivated to continue. The stress experienced in this situation is a function of the volume of exercise performed, rather than the intensity, with central fatigue (in cycling) and muscle damage (in running) being the primary outcomes of stress.
In ‘heavy’ exercise (exercise performed above the lactate threshold but below the maximal steady state or critical power), a steady state is delayed by 10-20 minutes, but metabolic parameters will eventually stabilise at oxygen uptake values higher than initially anticipated. In the heavy domain, volume and intensity combine to impose the stress. Together, these serve to draw down muscle fuel stores (chiefly glycogen) and provide a thermal load which may exceed the capacity to dissipate heat in hot and humid environments. The heavy domain has, therefore, been the basis of many classic studies on carbohydrate and fluid intake in the exercising human (Burnley & Jones, 2016). Above the maximal steady state or critical power (so-called severe-intensity exercise), none of the metabolic parameters associated with exercise can stabilise: oxygen uptake, phosphorylcreatine, inorganic phosphate, and pH all change progressively until exercise is terminated at task failure. In exercise engaging a large muscle mass, task failure follows the attainment of maximal oxygen uptake. The stress of severe-intensity exercise is derived primarily from its intensity, since task failure usually occurs in considerably less than 30 minutes. Severe-intensity exercise is, therefore, representative of the stress placed on humans exercising during endurance activities of ~2-30 minutes duration.
Overall, the above scheme is relevant to stress placed on the cardiorespiratory system and muscular systems during many endurance activities, but similar intensity/duration descriptors have been developed for exercise used to express and develop strength and maximal speed. These are usually expressed as some percentage of maximal static or dynamic muscle function (e.g. the ‘1 repetition maximum’). Both forms of exercise intensity quantification are used to structure exercise training programmes in order to provide sufficient stress to elicit the desired effect whilst avoiding injury (see below).
Exercise stress testing
By progressively increasing exercise intensity (treadmill speed or cycling power output, for example), it is possible to evaluate the body’s response to stress across the aerobic intensity spectrum described above. This is the basis of the classic incremental exercise test, variants of which are used throughout the world in cardiac or respiratory stress testing and the physiological evaluation of athletes. Of course, it is impossible to perform exercise that only stresses the heart or lungs, and so terms like ‘cardiac stress testing’ are something of a misnomer. Nevertheless, incremental exercise testing with electrocardiography, echocardiography, and pulmonary gas exchange measurements can reveal an array of cardiopulmonary problems, since few interventions provide the potent yet controlled stress that exercise does. That stress produces some unique signatures of cardiorespiratory dysfunction. For example, right-to-left atrial shunt caused by a patent foramen ovale in patients with pulmonary vascular disease is easily observed during incremental exercise (Wasserman et al., 1999), and indeed shunting may only occur during exercise. The same testing can quantify the effects of treatment, allowing the clinician to decide whether or not lung transplantation is required.
Stress testing in athletes is required for setting training intensity zones and, where relevant, for performance prediction. In these cases, the stress placed on the body must be as specific as possible in order to yield relevant performance information.
As a result, ergometers are not just mode-specific but are usually bespoke in the sense that they utilise as much of the athlete’s personal equipment as possible. The advent of highly accurate force and power measuring devices has all but eliminated the need for regular laboratory assessments of fitness in elite cycling, and the development of wearable sensors in other sports is likely to achieve the same end. Even though the modern-day sports physiologist’s laboratory is becoming increasingly portable, measurements in the field seldom match the precision and control of the laboratory environment. Translation of findings from the field to the laboratory and vice versa will remain a major area of the sports physiologist’s work for the foreseeable future.
Chronic responses to exercise stress: training
The well-documented effects of physical training are a manifestation of the chronic adaptation to exercise stress. Selye (1936) considered muscular exercise to be a form of his ‘general adaptation syndrome’, although in later writings he made a distinction between muscular adaptations to exercise and the non-specific stress reaction he was describing. A key feature of exercise stress is that adaptations to it are positive in all but the most extreme cases (overreaching or overtraining). In other words, application of training stress that does not cause injury will maintain or improve physiological function, in contrast to the so-called ‘exhaustion phase’ of the general adaptation syndrome.
Although the positive effect of training on performance capability and physiological parameters such as the maximal oxygen uptake have been known for decades, it is only relatively recently that changes in the morphological components that determine such parameters, and the molecular bases of such changes, have been characterised. Even so, only the molecular basis of ‘strength’ or ‘resistance’ training and ‘endurance’ training have so far been convincingly differentiated. Even in these cases our understanding of the processes leading to training adaptations is far from complete. To observe the clearest distinction between strength and endurance training usually requires a sample of trained participants. This is because even relatively low-intensity exercise in untrained individuals results in both mitochondrial and myofibrillar protein synthesis (see Coffey & Hawley [2017] for review). Additionally, for logistical reasons physical training studies rarely last more than 8-12 weeks, which severely limits our ability to provide evidence-based advice to athletes who have been training for many years.
The development of an ‘endurance’ phenotype, as one might expect, seems to require a high energetic demand from oxidative processes, but does not require the production of high forces. As detailed above, such high-intensity aerobic exercise produces a considerable metabolic stress. This results in AMP-activated protein kinase phosphorylation and subsequent transcription of mitochondrial proteins, leading to increased mitochondrial volume and aerobic enzyme activity. At the same time, angiogenesis (initiated, in part, by the expression of vascular endothelial growth factor [VEGF]) results in a greater capillary density. There is evidence that almost every step in the O2 conductance pathway beyond the alveoli can be enhanced by endurance training. These adaptations lead to a profound reduction in the stress experienced at a given absolute intensity. As a result, for a given level of effort, exercise performance improves (Fig.2).
Subjecting the muscle to very high forces in resistance training results in a completely different stressor and thus a completely different response to endurance exercise. Studies showing gains in maximal strength without measurable muscle hypertrophy lead to the inference that the strength gain has a neural origin. The behaviour of the motor unit pool in response to training is, however, extremely challenging to measure. It is only very recently, for example, that individual motor unit behaviour has been tracked longitudinally (Martinez-Valdes et al., 2017), and these data suggest that different forms of training exert differential effects on motor unit firing rate. In the fed state, muscle hypertrophy undoubtedly occurs in response to resistance training, and a number of factors that respond to mechanical stress are likely involved in signalling cascades leading to the deposition of new contractile proteins. The ‘downstream’ events resulting in protein synthesis have been thoroughly investigated in the last 10-15 years. Briefly, hypertrophy appears to be the result of the upregulation of protein synthesis primarily by the mechanistic target of the rapamycin (mTOR) pathway. In addition, the effects of exercise on connective tissue and bone remodelling to support high-intensity muscular contraction should not be forgotten. These responses may play an important role in healthy ageing by the avoidance of frailty in later life. Indeed, it is because exercise affects so many organ systems that its role in primary preventative healthcare is currently of interest.
Detraining: use it or lose it
The final twist in the story of exercise-induced stress concerns the effects of reducing or removing that stress. Many of the training-induced adaptations mentioned above begin to recede when training is ceased. If exercise intensity is maintained, however, so are the training-induced gains, a fact that athletes use in preparation for competition (the so-called ‘taper’ phase). Complete cessation of exercise leads to a process of detraining, in which muscle size, and mitochondrial and capillary densities decrease. These changes occur within days or weeks of stopping training. Central cardiovascular adaptations, such as left ventricular volume and blood volume revert to their pre-training values over a period of months. These detraining-induced losses can explained, in part, by the fact that these adaptations (such as the gain in lean tissue mass) are energetically expensive to maintain. In this context, detraining would appear to be a cost-saving process. As noted above, however, small amounts of relatively intense exercise appear capable of maintaining the gains accrued by previous training.
Conclusions
The acute physiological response to exercise stress is dominated by muscular, cardiovascular, and respiratory feedforward and feedback mechanisms, all of which attempt to limit the disruption to homeostasis produced by muscular contractions. The disruption to homeostasis results in fatigue, and produces the metabolic and/or mechanical signals that initiate the synthesis of new proteins. Whether these new proteins build new mitochondria or new contractile elements depends upon the nature of the exercise stress. As a result, exercise stress tries to break us, but its repeated application ultimately shapes us.
Here is the interesting part: "Telomere length increased significantly with sport intensity, especially in the younger group. HI had longer telomeres and higher levels of pro- and anti-inflammatory cytokines, suggesting less aging in HI compared to low and moderate counterparts in association with heightened immune response."
Increasing anti-inflammatory cytokines is so important in fighting disease and viruses.
Here is a good article about how those that went to European health clubs showed a low incident of Covid 19. I saw another study done Nordic countries that showed the same thing.